sytoskeletonia koostuu dynaamisesti vaihdettavasta verkosta, joka koostuu kolmesta erilaisesta proteiinilangasta solujen sytoplasmassa.
Ne antavat soluille ja organisatorisille solunsisäisille rakenteille kuten organelleille ja vesikkeleille rakenteen, stabiilisuuden ja luontaisen liikkuvuuden (liikkuvuuden). Jotkut filamentit ulkonevat solusta solun liikkuvuuden tai vieraiden kappaleiden suunnatun kuljetuksen tukemiseksi silikoiden tai silmujen muodossa.
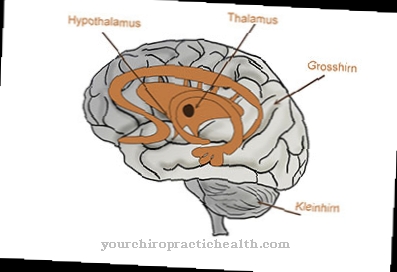
Mikä on sytoskeleton?
Ihmisen solujen sytoskeleton koostuu kolmesta eri luokasta proteiinilankoja. Mikrofilamentit (aktiinifilamentit), joiden halkaisija on 7 - 8 nanometriä ja jotka koostuvat pääasiassa aktiiniproteiineista, toimivat solun ulkoisen muodon ja koko solun liikkuvuuden sekä solunsisäisten rakenteiden stabiloimiseksi.
Lihassoluissa aktiinilangat mahdollistavat lihaksen supistumisen koordinoidulla tavalla. Välilangat, jotka ovat noin 10 nanometriä paksuja, tarjoavat myös kennolle mekaanisen lujuuden ja rakenteen. He eivät ole mukana solumotilassa. Välikefilamentit koostuvat erilaisista proteiineista ja proteiinien dimeereistä, jotka yhdistyvät muodostaen kimppuja, jotka on haavattu kuten köydet (tonofibrillit) ja jotka ovat erittäin repäisykestäviä rakenteita. Välilangat voidaan jakaa vähintään kuuteen erityyppiseen tyyppiin, joilla on eri tehtävät.
Kolmas filamenttiluokka koostuu pienistä putkista, mikroputkista, joiden ulkohalkaisija on 25 nanometriä. Ne koostuvat tubuliinidimeerien polymeereistä ja ovat ensisijaisesti vastuussa kaikentyyppisistä solunsisäisistä liikkuvuuksista ja solujen itsensä liikkuvuudesta. Solujen oman liikkuvuuden tukemiseksi silikaan tai flagellan muodossa olevat mikrotubulukset voivat muodostaa soluprosesseja, jotka työntyvät ulos solusta. Mikrotubulusten verkko on pääosin järjestetty sentromeeristä ja siihen kohdistuvat erittäin dynaamiset muutokset.
Anatomia ja rakenne
Aineiden ryhmät mikrofilamentit, välifilamentit (IF) ja mikrotubulit (MT), jotka kaikki kolme on osoitettu sytoskeletonille, ovat lähes kaikkialla läsnä sytoplasmassa ja myös solutumassa.
Ihmisten mikro- tai aktiini-filamenttien perusrakenteet koostuvat 6 isoformiaktiiniproteiinista, joista kukin eroaa vain muutamalla aminohapolla. Monomeerinen aktiiniproteiini (G-aktiini) sitoo nukleotidi ATP: n ja muodostaa aktiinimonomeerien pitkiä molekyyliketjuja jakamalla fosfaattiryhmän, joista kaksi yhdistyy muodostaen kierteisiä aktiinifilamentteja. Aktiinilangat sileissä ja nauhoitetuissa lihaksissa, sydänlihaksissa ja ei-lihaksiset aktiinilangat eroavat toisistaan hieman. Aktiinilankojen muodostuminen ja hajoaminen ovat erittäin dynaamisten prosessien alaisia, ja ne mukautuvat vaatimuksiin.
Välituotefilamentit koostuvat erilaisista rakenneproteiineista, ja niiden vetolujuus on korkea, joiden poikkileikkaus on noin 8 - 11 nanometriä. Välifilamentit jaetaan viiteen luokkaan: happamat keratiinit, emäksiset keratiinit, desmiinityyppi, neurofilamentit ja laminaattityypit. Vaikka keratiinit esiintyvät epiteelisoluissa, desmiini-tyyppiset filamentit löytyvät sileiden ja juosteisten lihasten lihassoluista sekä sydänlihassoluista. Neurofilamentit, joita on käytännössä kaikissa hermosoluissa, koostuvat proteiineista, kuten Internexin, Nestin, NF-L, NF-M ja muut. Laminaattityyppiset väliefilamentit löytyvät kaikista karyoplasman ydinmembraanin solunytimistä.
Toiminto ja tehtävät
Sytoskeleton toiminta ja tehtävät eivät ole mitenkään rajoittuneet solujen rakenteelliseen muotoon ja stabiilisuuteen. Mikrofilamentit, jotka sijaitsevat pääasiassa verkkomaisissa rakenteissa suoraan plasmakalvolla, stabiloivat solujen ulkoisen muodon. Mutta ne muodostavat myös kalvon ulkonemat, kuten pseudopodia. Moottoriproteiinit, joista lihassolujen mikrofilamentit rakentuvat, varmistavat lihaksille tarvittavat supistukset.
Erittäin korkeat vetolujuudet väliffilamentit ovat erittäin tärkeitä kennojen mekaaniselle lujuudelle. Heillä on myös joukko muita toimintoja. Epiteelisolujen keratiinilangat yhdistetään epäsuorasti mekaanisesti toisiinsa desmosomien kautta siten, että ihokudos saa kaksiulotteisen, matriisimaisen lujuuden. IF: t on kytketty muihin sytoskeleton aineryhmiin filamenttivälitteisillä välituoteproteiineilla (IFAP), mikä varmistaa tietyn tiedonvaihdon ja vastaavan kudoksen mekaanisen lujuuden. Tämä luo järjestettyjä rakenteita sytoskeleton sisällä. Entsyymit, kuten kinaasit ja fosfataasit, varmistavat verkkojen rakentamisen, rakenneuudistuksen ja hajoamisen nopeasti.
Erityyppiset neurofilamentit stabiloivat hermokudosta. Laminaatit säätelevät solukalvon hajoamista solunjakautumisen ja sitä seuraavan jälleenrakennuksen aikana. Mikrotubulukset ovat vastuussa tehtävistä, kuten organelien ja rakkuloiden kuljetuksen kontrolloimisesta solussa ja kromosomien järjestämisestä mitoosin aikana. Soluissa, joissa mikrotubuluksissa kehittyy mikrovilli, silia, flagella tai flagella, MT: t myös varmistavat koko solun liikkuvuuden tai huolehtivat liman tai sellaisten vieraiden kappaleiden poistamisesta. B. henkitorvessa ja ulkoisessa korvakanavassa.
Löydät lääkkeesi täältä
Memory Muistihäiriöitä ja unohduksia ehkäisevät lääkkeetsairaudet
Häiriöt sytoskeleton aineenvaihdunnassa voivat johtua joko geneettisistä virheistä tai ulkoisesti toimitetuista toksiineista. Yksi yleisimmistä perinnöllisistä sairauksista, jotka liittyvät lihasten membraaniproteiinin synteesin häiriöihin, on Duchennen lihasdystrofia.
Geneettinen vika estää dystrofiinin, rakenteellisen proteiinin, muodostumisen, jota tarvitaan nauhoitettujen luustolihasten lihaskuituihin. Tauti esiintyy varhaislapsuudessa etenevällä tavalla. Mutatoituneilla keratiineilla voi olla myös vakavia vaikutuksia. Ichtioosi, ns. Kalan hilsetauti, johtaa hyperkeratoosiin, epätasapainoon ihon hiutaleiden muodostumisen ja kuorinnan välillä, johtuen yhdestä tai useammasta geenivirheestä kromosomissa 12. Ikytoosi on yleisin perinnöllinen ihosairaus ja vaatii intensiivistä terapiaa, joka voi kuitenkin vain lievittää oireita.
Muut geneettiset viat, jotka johtavat hermosäikeiden aineenvaihdunnan häiriöihin, aiheuttavat z: n. B. amyotrofinen lateraaliskleroosi (ALS). Jotkut tunnetut mykotoksiinit (sienimyrkyt), kuten muotista ja kärpäse-agarista peräisin olevat, häiritsevät aktiinilankojen metaboliaa. Kolkisiiniä, syksyn krookuksen toksiinia, ja taksolia, jota saadaan marjakuusta, käytetään erityisesti kasvainhoitoon. Ne puuttuvat mikrotubulusten aineenvaihduntaan.


.jpg)


.jpg)